 Ядро (лат. nucleus) — клітинна органела, знайдена у більшості клітин еукаріотів і містить ядерні гени, які складають більшу частину генетичного матеріалу. Ядро має дві первинні функції: керування хімічними реакціями в межах цитоплазми і збереження інформації, потрібної для поділу клітини. Окрім клітинного геному, ядро містить певні білки, які регулюють зчитування генетичної інформації.
Ядро (лат. nucleus) — клітинна органела, знайдена у більшості клітин еукаріотів і містить ядерні гени, які складають більшу частину генетичного матеріалу. Ядро має дві первинні функції: керування хімічними реакціями в межах цитоплазми і збереження інформації, потрібної для поділу клітини. Окрім клітинного геному, ядро містить певні білки, які регулюють зчитування генетичної інформації.
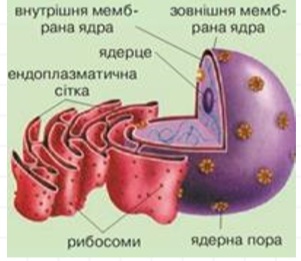
Ядро звичайно має розмір 8-25 мікрометрів у діаметрі. Воно оточено подвійною мембраною, яка називається ядерною оболонкою. Крізь внутрішню і зовнішню мембрани на деяких інтервалах проходять ядерні пори. Ядерна оболонка регулює і полегшує транспорт між ядром і цитоплазмою, відокремлюючи хімічні реакції, що відбуваються в цитоплазмі, від реакцій, що трапляються в межах ядра. Зовнішня мембрана безперервна з гранулярним ендоплазматичним ретикулумом і може мати зв’язані рибосоми. Простір між двома мембранами (який називається «перинуклеарним простором») безперервний і пов’язаний з ендоплазматичною сіткою. Ядерна сторона ядерної оболонки оточена мережею проміжних філаментів, яка називається ядерною ламіною. Внутрішня частина ядра містить одне або декілька ядерець, оточених матриксом, який називається нуклеоплазмою. Нуклеоплазма (каріолімфа, ядерний сік, каріоплазма) — гелеподібна рідина (подібна у цьому відношенні до цитоплазми), в якій розчинені багато речовин. Ці речовини включають нуклеотид-трифосфати, сигнальні молекули, ДНК, РНК та білки (ензими та філаменти).
Ядро та ендомембранна система.
ХРОМАТИН.
Генетичний матеріал присутній в ядрі у вигляді хроматину, або комплексу білка і ДНК. Є два види хроматину: еухроматин і гетерохроматин.

Еухроматин (синнє на малюнку)— менш компактна форма ДНК. Області ДНК, які знаходяться у формі еухроматину містять гени, які часто зчитуються клітиною.
У гетерохроматині (червоне на малюнку) ДНК більш компактно упакована. Області ДНК, які знаходяться у формі гетерохроматину містять гени, які не зчитуються клітиною на даній стадії (цей вид гетерохроматину відомий як факультативний гетерохроматин) або є областями, які складають теломери і центромери хромосом (цей вид гетерохроматину відомий як конструктивний гетерохроматин). У багатоклітинних організмах, клітини надзвичайно спеціалізовані, щоб виконувати специфічні функції, тому різні набори генів потрібні і зчитуються. Ядерце — щільна структура в ядрі, де збираються елементи рибосом. У ядрі може бути одне або декілька ядерець.
Хроматином називають молекули хромосомної ДНК в комплексі зі специфічними білками, необхідними для здійснення цих процесів. Основну масу складають «білки зберігання», так звані гістони. З цих білків побудовані нуклеосоми — структури, на які намотані нитки молекул ДНК. Нуклеосоми розташовуються досить регулярно, так що утворюється структура яка нагадує намисто. Нуклеосома складається з білків чотирьох типів: H2A, H2B, H3 і H4. В одну нуклеосому входять по два білка кожного типу — всього вісім білків. Гістон H1, більший ніж інші гістони, зв’язується з ДНК в місці її входу на нуклеосому. Нуклеосома разом з H1 називається хроматосомою. Нитка ДНК з нуклеосомами утворює нерегулярну соленоїд-подібну структуру товщиною близько 30 нанометрів, так звану 30 нм фібрилу. Подальша упаковка цієї фібрили може мати різну щільність. Як зазначалося, якщо хроматин упакований щільно, його називають конденсованим або гетерохроматином, його можна побачити під мікроскопом. У інтерфазі гетерохроматин звичайно розташовується по периферії ядра (пристінковий гетерохроматин). Повна конденсація хромосом відбувається перед поділом клітини. Якщо хроматин упакований нещільно, його називають еухроматином або інтерхроматином. Цей вид хроматину набагато менш щільний при спостереженні під мікроскопом і зазвичай характеризується наявністю транскрипційної активності. Щільність упаковки хроматину є частиною епігенетичного контролю експресії (вияву) генів і частково визначається модифікаціями гістонових хвостів — ацетилюванням та деацетилюванням і метилюванням.
З ІСТОРІЇ.
В 1831 році англійський природознавець Роберт Браун вивчав різні види рослин, зразки яких він зібрав під час подорожі до Австралії. Браун був дуже уважним до деталей, а клітини рослин особливо цікавили його. Розглядаючи їх під мікроскопом, він побачив дещо цікаве: кожна клітина містила круглий і непрозорий елемент. Він назвав його ядром. Дізнавшись про спостереження Брауна, німецький фізіолог Теодор Шванн почав шукати подібні елементи в клітинах пуголовків і знайшов. Кожна клітина містила ядро. Це був революційний прорив — свідчення того, що всі види життя пов’язані між собою.

В одній із книг Шванн описав різні типи клітин, взяті від різноманітних організмів і визначив їх за фактом наявності ядра. Усвідомлення того, що є елемент спільний для всіх організмів, не тільки для рослин, а й для тварин, поєднало рослинне і тваринне царство у щось спільне, щось, що мало однакові риси. Німецький біолог Хаммерлінг В 1935 році продемонстрував значення ядра як сховища генетичного матеріалу в експериментах на одноклітинній морській водорості ацетабулярії.
ХРОМОСОМИ
Хромосоми помітні тільки в момент поділу. З метою вивчення хромосом застосовують колхіцин, який гальмує стадію анафази, в результаті чого хроматиди не розходяться до полюсів клітини і мають вигляд Х-образних фігур.
Хромосома-— це велика молекулярна структура, де міститься близько 90 % ДНК клітини. Кожна хромосома містить дуже довгий безперервний полімеризований ланцюг ДНК (єдину ДНК-молекулу), що містить гени, регуляторні елементи та проміжні нуклеотидні послідовності. Слово «хромосома» походить від грецьких слів «хрома» — колір та «сома» — тіло. Хромосоми еукаріот складаються з лінійної макромолекули ДНК, що намотана на специфічні білки-гістони, формуючи матеріал під назвою «хроматин». В клітинах прокаріот звичайно міститься єдина хромосома, яка, на відміну від еукаріот, є кільцевою та позбавленою гістонів. Втім, це правило не є абсолютним: існують бактерії з більше, ніж одною хромосомою; у деяких бактерій хромосоми є лінійними; у кількох видів архей виявлені специфічні гістони. Хромосоми можуть перебувати в двох структурно-функціональних станах: в конденсованому (спіралізованому) та деконденсованому (деспіралезованому). В інтерфазі хромосоми живої клітини невидимі, спостерігати можна лише гранули хроматину, бо в цей період хромосоми частково або повністю деконденсовані. Це є їхнім робочим станом, бо в більш дифузномухроматині активніші процеси синтезу. Під час мітотичного поділу клітини, коли відбувається конденсація хроматину, хромосоми добре помітні у світловий мікроскоп. Мітотичні хромосоми складаються із двох ідентичних молекул ДНК (попередньо ДНК подвоюється, реплікується). Фактично, хроматин-це сукупність хромосом, що складаються кожна з однієї неконденсованої молекули ДНК. Хромосоми складаються з двох молекул ДНК, що поєднуються перетяжкою-центромерою.

Будова хромосом, залежно від розташування центромери
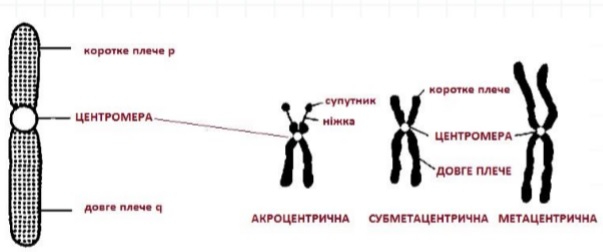
- ЦЕНТРОМЕРА (от центр + греч. meros — частина) — спеціалізована ділянка ДНК, в районі якої в стадії профази та метафази поділу клітини з’єднуються дві хроматиди, що утворилися в результаті дуплікації (подвоєння) хромосоми.
- ХРОМАТИДА (от греч. chroma – колір, фарба + eidos – вид) — частина хромосоми від момента її дупликації (подвоєння) до розділення на дві дочірні в анафазі, являє собою нитку молекули ДНК що з’єднана білками-гістонами.
- ТЕЛОМЕРА (від др.-греч. τέλος – кінець і μέρος – частина) – кінцеві ділянки хромосом. Теломерні ділянки хромосом характеризуються відсутністю здатності до з’єднання з іншими хромосомами або їх фрагментами і виконують захисну функцію. Термін «теломера» запропонував Г. Меллер в 1932 р. у 2009 році присуджена Нобелевська премія з відкриття значення теломер у процесах старіння. З віком теломери хромосом зменшуються.
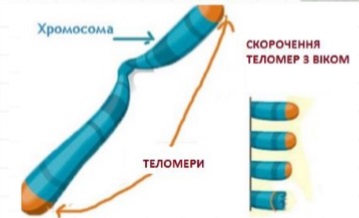
- Хроматиди утворюються в результаті дуплікації хромосоми у процесі поділу клітини.
- Хромосоми є у ядрах всіх клітин.
- Кожна хромосома містить спадкові інструкції – гени.
Стадії компактизації (укладання, конденсації) хроматину
Хромати́н — комплекс молекул ДНК та специфічних білків, що складає хромосоми.
Гістон– великий клас ядерних білків, що виконують дві основні функції: вони беруть участь в упаковці ниток ДНК в ядрі і в епігенетичною регуляції таких ядерних процесів, як транскрипція, реплікація і репарація.
Нуклеосома– структурна частина хроматину, утворена ділянкою нитки ДНК намотаною на серцевину із основних білків-гістонів, має діаметр ~11 нм.
Соленоїд– ланцюг нуклеосом, що утворюється на другому рівні компактизації хроматину згорнутий в симетрично, що містить нуклеомери.
- Перший рівень компактизації ДНК – нуклеосомний. Якщо піддати дії нуклеази хроматин, то він і ДНК, піддаються розпаду на регулярно повторювані структури. Після нуклеазної обробки з хроматину шляхом центрифугування виділяють фракцію частинок зі швидкістю седиментації (осадження)11S. Частинки 11S містять ДНК близько 200 нуклеотидних пар і вісім гістонів. Така складна нуклеопротеїдна частинка отримала назву нуклеосоми. У ній гістони утворюють білкову основусерцевину, по поверхні якої розташовується ДНК. ДНК утворює ділянку, що не пов’язана з білками серцевини – лінкер, який, з’єднуючи дві сусідні нуклеосоми, переходить в ДНК наступної нуклеосоми. Вони утворюють «намистини», глобулярні утвори близько 10 нм, що сидять один за одним на витягнутих молекулах ДНК.
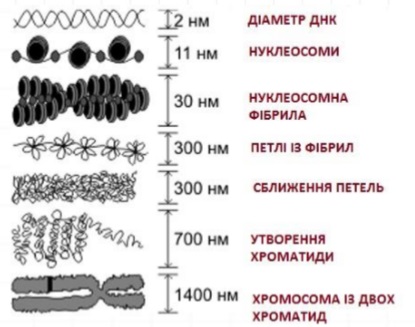
- Другий рівень компактизації-30 нм фібрила. Перше, нуклеосомний рівень компактизації хроматину відіграє регуляторну та структурну роль, забезпечуючи щільність упаковки ДНК в 6-7 разів. У мітотичних хромосомах і в інтерфазних ядрах виявляються фібрили хроматину з діаметром 25-30 нм. Виділяють соленоїдний тип укладання нуклеосом: нитка щільно упакованих нуклеосом діаметром 10 нм утворює витки з кроком спіралі близько 10 нм. На один виток такої суперспіралі припадає 6-7 нуклеосом. В результаті такої упаковки виникає фібрила спірального типу з центральною порожниною. Хроматин у складі ядер має 25-нм фібрили, яка складається з зближених глобул того ж розміру – нуклеомерів. Ці нуклеомери називають зверх бусинами («супербіди»). Основна фібрила хроматину діаметром 25 нм являє собою лінійне чергування нуклеомерів уздовж компактизованної молекули ДНК. У складі нуклеомера утворюються два витки нуклеосомної фібрили, по 4 нуклеосоми в кожному. Нуклеомерний рівень укладання хроматину забезпечує 40кратне ущільнення ДНК. Нуклесомний і нуклеомерний (супербідний) рівні компактизації ДНК хроматину здійснюються за рахунок гістонових білків.
- Петльові домени ДНК – третій рівень структурної організації хроматину. У вищих рівнях організації хроматину специфічні білки зв’язуються з особливими ділянками ДНК, яка в місцях зв’язування утворює великі петлі, або домени. У деяких місцях є згустки конденсованого хроматину, розеткоподібні утворення, що складаються з багатьох петель 30 нмфібрил, що з’єднуються в щільному центрі. Середній розмір розеток досягає 100-150 нм. Розетки фібрил хроматинуХромомери. Кожен хромомер складається з декількох петель, що містять нуклеосоми, які пов’язані в одному центрі. Хромомери пов’язані один з одним ділянками нуклеосомного хроматину. Така петельно-доменна структура хроматину забезпечує структурну компактизацію хроматину і організовує функціональні одиниці хромосом – реплікони і транскрибуючі гени.
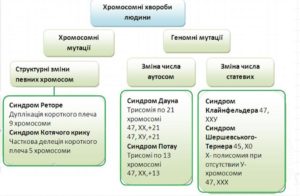
ГОМОЛОГІЧНІ ХРОМОСОМИ Від грец. Гомос – однаковий Гомологічні хромосоми – парні хромосоми, одинакові за формою, размірами та за набором генів.
ДИПЛОЇДНИЙ НАБІР. У клітинах тіла двостатевих тварин і рослин кожна хромосома представлена двома гомологічними хромосомами, що відбуваються одна від материнського, а інша від батьківського організму. Такий набір хромосом називають диплоїдним (подвійним)
ГАПЛОЇДНИЙ НАБІР ХРОМОСОМ. Статеві клітини, що утворилися в результаті мейозу, містять тільки одну з двох гомологічних хромосом. Цей набір хромосом називають гаплоїдним (одинарним).
КАРІОТИП – це сукупність числа, розмірів та морфології мітотичних хромосом (у людини 46). ГЕНОМ – гаплоїдний хромосомний набір; сукупність генів, локалізованих в одиночному наборі хромосом даного організму. Термін запропонований в 1920 німецьким біологом Г. Винклером.
ГЕНОТИП – сукупність генів даного організму, яка, на відміну від понять геному і генофонду, характеризує особину, а не вид (також генотип відрізняється від генома, оскільки в поняття “геном” включають і некодуючі послідовності, що не входить у поняття “генотип”). Разом з чинниками зовнішнього середовища визначає фенотип організму.
ГЕНОФОНД – поняття з популяційної генетики, що описує сукупність всіх генних варіацій ( алелей) певної популяції. Популяція володіє всіма своїми алелями для оптимального пристосування до навколишнього середовища. Можна також говорити про єдиний генофонд виду, так як між різними популяціями виду відбувається обмін генами.
ФЕНОТИП – сукупність характеристик, властивих індивіду на певній стадії розвитку. Фенотип формується на основі генотипу, опосередкованого низкою зовнішніх факторів. У диплоїдних організмів у фенотипі проявляються домінантні гени. ГЕН-носій спадкової інформації, відповідає за синтез білка, вияв певної ознаки. Гени лежать в лінійному порядку (в ряд, один за одним)на протязі всієї хромосоми.
ПОРУШЕННЯ СТРУКТУРИ ХРОМОСОМ
Відбувається в результаті спонтанних або спровокованих змін: Генні (точкові) мутації (зміни на молекулярному рівні)
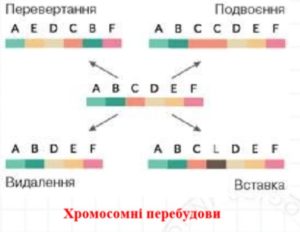
Аберрації (хромосомні перебудови, розрізняються при збільшенні на світловому мікроскопі).
Хромосомні перебудови бувають:
Внутрішньохромосомні – структурні зміни в межах однієї хромосоми:
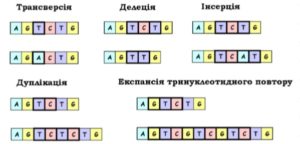
- Дефішенсії – втрата хромосомою її кінцевої ділянки, термінальна делеція.
- Делеції – втрата хромосомою її внутрішньої ділянки, інтерстиціальні делеції. Може бути наслідком розриву хромосоми або результатом нерівного кроссинговеру. Наприклад, делеція білка CCR5-дельта32 приводить до неприйнятності її носія до ВІЛ. Зараз до ВІЛ виявляє стійкість в средньому 10 % європейців, однак в Скандинавії ця доля досягає 14-15 %. У фіннів і росіян доля стійких людей ще вище — 16 %, а в Сардинії — всього 4 %.
- Дуплікації лат. duplicatio– подовження однієї хроматини за рахунок вбудування іншої, сестринської.
- Інверсії – Це зміни структури хромосоми, викликані поворотом на 180° однієї із внутрішнніх її ділянок. Подібна перебудова – це наслідок двух одночасних розривів в одній хромосом.
Міжхромосомні- структурні зміни, що відбуваються між двома та більшою кількістю хромосом.
- Транспозиція –переміщення генів у межах однієї чи декількох хромосом. Займає проміжне положення між внутрішньо і між хромосомними перебудовами.
- Транслокація – відбувається обмін ділянками негомологічних хромосом, але загальна кількість генів не змінюється. Різні транслокації приводять до розвитку лімфом, сарком, захворюванню лейкемією, шизофренією.

Моногенні хвороби (зміна у структурі гена)
Аутосомно-домінантніми –полідактелія (шестипалість), хорея Гентінгтона
Аутосомно-рецесивними – фенилкетанурія, альбінізм
Х-зчеплені рецесивні – гемофілія, дальтонізм
У- зчеплені оволосеніння вушної раковини
Аномалії аутосом.
Моносомія по будь-якій з аутосом зазвичай призводить до внутрішньоутробної загибелі плоду.
Моносомія – найчастіший варіант хромосомної аномалії при мимовільних абортах. Трисомії по аутосомам характеризуються меншою летальністю, але викликають важкі вади розвитку у дітей, народжених живими. У хворих з мозаїцизмом клінічна картина менш виражена, а виживаність вище, ніж при повній формі трисомії. За допомогою диференціального фарбування хромосом було встановлено, що чисельні зміни в системі аутосом часто супроводжуються невеликими делеціями і дуплікаціями. Якщо у пробанда виявлені чисельні або структурні аномалії хромосом, досліджують каріотип батьків для виключення збалансованих перебудов (це необхідно для визначення повторного ризику народження хворої дитини)
ХВОРОБИ ЇЗ СПАДКОВОЮ СХИЛЬНІСТЮ (МУЛЬТИФАКТОРІАЛЬНІ АБО ПОЛІГЕННІ)
1. Характеризуються генетичною гетерогенністю та клінічним поліморфізмом;
2. Реалізується при певній відповідностігенетичних та середовищ них факторів;
3. (цукровий діабет, хвороба Альцгеймера, атеросклероз, гіпертонічна хвороба, шизофренія