










ЕНДОПЛАЗМАТИЧНА СІТКА АБО РЕТИКУЛЮМ
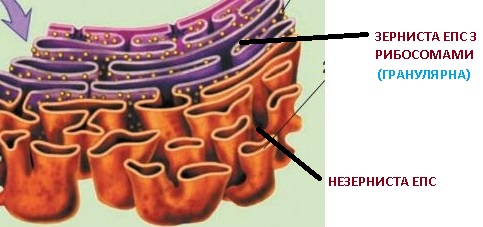
Ендоплазматична сітка становить собою систему порожнин у вигляді мікроскопічних канальців та їхніх потовщень (так званих цистерн). Діаметр канальців становить 50–100 нм, а цистерн – до 1000 нм і більше. Вони обмежені мембраною та сполучаються між собою. Розріз няють два різновиди ендоплазматичної сітки: зернисту та незернисту. Обидва різновиди ендоплазматичної сітки мають тісні просторові зв’язки; зокрема, їхні мембрани можуть безпосередньо переходити одна в іншу.
Одна з основних функцій зернистої ендоплазматичної сітки – забезпечення транспорту білків по клітині.Частина синтезованих у клітині білків використовується для її власних потреб, а частина виводиться за межі клітини. Білки синтезуються за участі рибосом, які можуть розташовуватися в цитозолі та на поверхні зернистої едоплазматичної сітки.
У її порожнинах білки набувають притаманної їм просторової конформації, до них можуть приєднуватися небілкові компоненти. Синтезовані білки використовуються для побудови плазматичної мембрани та зовнішньої мембрани оболонки ядра в період між поділами клітини.
На мембранах незернистої ендоплазматичної сітки синтезуються ліпіди, вуглеводи, певні гормони, що можуть накопичуватися в її порожнинах. У цих порожнинах (наприклад, у клітинах печінки) накопичуються і знешкоджуються деякі отруйні сполуки.
Крім того, порожнини сітки м’язових клітин, нейронів, залозистого епітелію слугують внутрішньоклітинним депо (місцем зберігання) йонів Кальцію.
Це має важливе значення для функціонування м’язових волоконець: під час їхнього скорочення концентрація йонів Кальцію різко підвищується, а під час розслаблення – знижується. Ендоплазматична сітка функціо-нально пов’язана з комплексом Гольджі.
КОМПЛЕКС ГОЛЬДЖІ
Так (названий так на честь італійського вченого К. Гольджі, який відкрив цю органелу) – одна з універсальних органел клітин еукаріотів. Її основною структурною одиницею слугує диктіосома – скупчення одномембранних пласких цистерн. Поруч з ними розташовані пухирці та канальці. До одного з полюсів кожної із цистерн постійно надходять пухирці, які відокремлюються від ендоплазматичної сітки і містять речовини, що там утворилися.
Зливаючись із цистернами комплексу Гольджі, ці пухирці віддають їм свій вміст. Функції комплексу Гольджі різноманітні. Насамперед у цій органелі накопичуються і певним чином змінюються деякі сполуки (наприклад, білки можуть сполучатися з вуглеводами або ліпідами). Речовини, які надійшли до цистерн комплексу Гольджі, сортуються за хімічним складом і призначенням. Відсортовані молекули переходять з одних цистерн до інших і згодом у ви-гляді пухирців, оточених мембраною, відокремлюються від цієї органели. У цистернах комплексу Гольджі синтезуються деякі полісахариди.
Відсортовані молекули переходять з одних цистерн до інших і згодом у ви-гляді пухирців, оточених мембраною, відокремлюються від цієї органели. У цистернах комплексу Гольджі синтезуються деякі полісахариди.
Вони можуть сполучатися з білками, що надійшли із зернистої ендоплаз-матичної сітки. У рослинних клітинах у комплексі Гольджі утворюються структурні компоненти клітинної стінки, а в клітинах членистоноги-хітиновмісної кутикули, яка утворює зовнішній скелет.
Завдяки комплексу Гольджі в голівці сперматозоїдів утворюється списо- або чашоподібний утвір –акросома (від грец. акрон – верхівка). Ця органела містить ферменти, які розчиняють оболонку яйцеклітини під час запліднення. Скоротливі вакуолі прісноводних одноклітинних тварин і водоростей також формуються з комплексу Гольджі. Ця органела бере участь і в побудові плазматичної мембрани. Комплекс Гольджі бере участь у формуванні лізосом, які у вигляді пухирців, оточених мембраною, відокремлюються від цієї органели.
ЛІЗОСОМИ
Від (грец. лізіс – розчинення) це – органели, які мають вигляд мікроскопічних оточених мембраною пухирців діаметром 100–180 нм.
Вони містять ферменти, здатні розщеплювати ті чи інші сполуки (білки, вуглеводи, ліпіди тощо). У клітині можуть бути різні види лізосом, які відрізняються особливостями будови і функціями.
Одні з лізосом, зливаючися з піноцитозними або фагоцитозними пухирцями, беруть участь у формуванні травних вакуоль.
При цьому активуються ферменти і вміст вакуолі перетравлюється.
Таким чином, лізосоми забезпечують процеси внутрішньоклітинного травлення. Лізосоми іншого типу беруть участь у перетравленні окремих частин клітини цілих клітин або їхніх груп. Вважають, що ці лізосоми знищують дефектні органели, ушкоджені або мертві клітини, руйнують личинкові органи комах, земноводних (наприклад, хвости та зябра пуголовків безхвостих амфібій) тощо. Лізосоми можуть наближатися до плазматичної мембрани та виводити свій вміст – ферменти – назовні. Наприклад, у гриба нейроспори у такий спосіб забезпечуються процеси позаклітинного травлення.
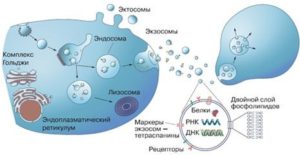 Везикулярний транспорт. Спочатку ці крихітні бульбашки- везикули розглядалися як резервуари для видалення надлишку клітинної цитоплазми і вважалися побічним продуктом життєдіяльності клітин. З моменту їх відкриття і до початку XXI століття ніхто ними особливо не цікавився. Потім з’ясувалося, що вони мають здатність регулювати імунні реакції організму. Коли ж у 2007 році в них були виявлені нуклеїнові кислоти – носії генетичної інформації, стало ясно, що це зовсім не сміттєві контейнери. Везикулярний транспорт відбувається за допомогою везикул -маленьких бульбашок, оточених мембраною, що містять різні речовини.
Везикулярний транспорт. Спочатку ці крихітні бульбашки- везикули розглядалися як резервуари для видалення надлишку клітинної цитоплазми і вважалися побічним продуктом життєдіяльності клітин. З моменту їх відкриття і до початку XXI століття ніхто ними особливо не цікавився. Потім з’ясувалося, що вони мають здатність регулювати імунні реакції організму. Коли ж у 2007 році в них були виявлені нуклеїнові кислоти – носії генетичної інформації, стало ясно, що це зовсім не сміттєві контейнери. Везикулярний транспорт відбувається за допомогою везикул -маленьких бульбашок, оточених мембраною, що містять різні речовини.
Як зазначається в наукових роботах лауреатів Нобелівської премії, кожна везикула містить у своїй мембрані білкові молекули- “поштові адреси”, що дозволяють доставляти її за призначенням – як всередині клітин, так і між клітинами.
ВАКУОЛІ
Від (лат. вакуус – порожній) – одномембранні органели, що міститься в деяких еукаріотичних клітинах, мають вигляд порожнин, оточених мембраною і заповнених рідиною і виконує різні функції (секреція, екскреція і зберігання запасних речовин, аутофагія, автоліз та ін.) Вакуолі і їх вміст розглядаються як відокремлений від цитоплазми компартмент. Розрізняють травні і скоротливі (пульсуючі) вакуолі, що регулюють осмотичний тиск і слугують для виведення з організму продуктів розпаду. Вакуолі особливо добре помітні в клітинах рослин: у багатьох зрілих клітинах рослин вони складають більше половини обсягу клітини. Одна з важливих функцій рослинних вакуолей – накопичення іонів і підтримка тургору (тургорного тиску). Вакуоль – це місце запасу води. Вакуолі розвиваються з цистерн ендоплазматичної сітки.
Мембрана, в якою оточена вакуоль, називається тонопласт. У вакуолях містяться органічні кислоти, вуглеводи, дубильні речовини, неорганічні речовини (нітрати, фосфати, хлориди), білки.
Розрізняють різні види вакуоль. Про утворення травних вакуоль, у яких перетравлюються сполуки і мікроорганізми, що надходять у клітину, ми вже згадували.
Вакуолі рослинних клітин утворюються з пухирців, які відокремлюються від ендоплазматичної сітки. Згодом дрібні вакуолі зливаються в більші, які можуть охоплювати майже весь об’єм цитоплазми. Вони заповнені клітинним соком – водним розчином органічних і неорганічних сполук. Вакуолі підтримують певний рівень внутрішньоклітинного тиску (тургору), забезпечуючи збереження форми клітин, містять запасні поживні речовини, кінцеві продукти обміну або пігменти. Червоні, сині, жовті тощо пігменти, розчинені у клітинному соку, зумовлюють забарвлення певних типів клітин і частин рослин у цілому (наприклад, плодів вишні, коренеплодів редису, пелюсток квітів тощо). Через мембрани вакуоль речовини переміщуються із цитозолю в їхню порожнину і навпаки.
У клітинах прісноводних одноклітинних тварин і водоростей є скоротливі вакуолі – вкриті мембраною пухирці, здатні змінювати об’єм, виводячи свій вміст назовні. Це пов’язано з тим, що в прісній воді концентрація солей значно нижча, ніж у цитоплазмі. Тому згідно з фізичними законами вода з навколишнього середовища надходить до клітини, підвищуючи тиск усередині неї. Надлишок рідини накопичується в скоротливій вакуолі. Стінка вакуолі стискається завдяки скоротливим білкам, які входять до її складу, і випорскує рідину назовні клітини. Таким чином, скоротливі вакуолі регулюють внутрішньоклітинний тиск, виводячи надлишок води з клітини і запобігаючи її руйнуванню. Крім того, ці органели забезпечують виведення деяких розчинних продуктів обміну речовин. Діяльність скоротливої вакуолі має значення й для газообміну, оскільки сприяє надходженню до клітини води з розчиненим киснем і виведенню вуглекислого газу. Найскладнішу будову скоротливих вакуоль мають інфузорії. Наприклад, в інфузорії- туфельки вакуоля складається зі скоротливого пухирця (резервуару), який відкривається назовні отвором – порою. У пухирець впадають довгі тоненькі радіальні канальці, до яких надходять водні розчини із цитоплазми. Спочатку скорочуються стінки канальців, проштовхуючи рідину до резервуару. Через певний час скорочується наповнений резервуар, викидаючи вміст через пору.
ПЕРОКСИСОМИ
Від грец. пери – навколо, лат. окси – Оксиген та грец. сома – тіло), або мікротільця – органели кулястої форми, оточені однією мембраною, діаметром близько 0,3–1,5 мкм. Пероксисоми виявлені в різних організмах: одноклітинних і багатоклітинних тварин, дріжджів, вищих рослин, водоростей. Ці органели часто розташовані поблизу мембран ендоплазматичної сітки, мітохондрій, пластид. У них містяться різноманітні ферменти, зокрема ті, що забезпечують перетво-рення жирів на вуглеводи або здатні розщеплювати токсичний для клітини гідроген пероксид Н 2 О 2 до кисню та води. У рослин пероксисоми беруть участь у процесах поглинання кисню під час світлової фази фотосинтезу.
Тривалість існування однієї пероксисоми незначна – лише 5–6 діб. Нові пероксисоми утворюються внаслідок поділу.
НЕМЕМБРАННІ ОРГАНЕЛИ
 РИБОСОМИ (ribosome) є немембранною органелою клітини, що складається з рРНК та рибосомних білків (протеїнів). Рибосома здійснює біосинтез білків транслюючи з мРНК поліпептидний ланцюг. Таким чином, рибосому можна вважати фабрикою, що виготовляє білки, базуючись на наявній генетичній інформації. В клітині дозрілі рибосоми знаходяться переважно в компартментах, для активного білкового синтезу. Вони можуть вільно плавати
РИБОСОМИ (ribosome) є немембранною органелою клітини, що складається з рРНК та рибосомних білків (протеїнів). Рибосома здійснює біосинтез білків транслюючи з мРНК поліпептидний ланцюг. Таким чином, рибосому можна вважати фабрикою, що виготовляє білки, базуючись на наявній генетичній інформації. В клітині дозрілі рибосоми знаходяться переважно в компартментах, для активного білкового синтезу. Вони можуть вільно плавати
в цитоплазмі або бути прикріпленими до цитоплазматичного боку мембран ендоплазматичного ретикулуму чи ядра. Субодиниці рибосом укладаються у ядерці.
КЛІТИННИЙ ЦЕНТР (центросома) – немембранна органела еукаріотичнчх клітин, що складається з центріолей. Уперше виявлена в 1888 році Теодором Бовері, який назвав її “особливим органом клітинного поділу”. У клітині, яка перебуває на початку інтерфази, ця органела знаходиться біля ядра. Вона виявлена в усіх клітинах тварин (за винятком яйцеклітин), у водоростей (вищі рослини його не мають) і в клітинах грибів.
Клітинний центр складається, як правило, з двох центріолей, розташованих під прямим кутом одна до одної. Перебувають вони в ділянці світлої цитоплазми (центросфери), від якої розходяться мікротрубочки.
Аномальне збільшення кількості центріолей характерне для багатьох ракових клітин. Кожна центріоля клітинного центру має вигляд порожнистого циліндра діаметром близько 0,15 мкм і довжиною 0,5 мкм. Стінки циліндра формує віночок з 9 груп мікротрубочок (по 3 мікротрубочки в групі, тобто з 9 триплетів), розміщених по колу. Наприкінці інтерфази біля кожної материнської центріолі утворюється дочірня, дещо коротша, розміщена перпендикулярно до материнської. Таким чином, перед мітозом клітина містить дві пари центріолей.
Активна роль клітинного центру виявляється при поділі клітини. Центріолі розходяться до полюсів клітини і організовують розміщення мікротрубочок у структуру, яка має назву веретена поділу.
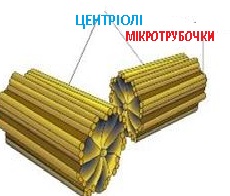 Центріолі беруть участь в утворенні мікротрубочок цитоскелета. Крім участі в поділі ядра, клітинний центр відіграє важливу роль у формуванні еукаріотичних джгутиків і війок. Його центріолі формують базальне тіло, що лежить в основі джгутиків.
Центріолі беруть участь в утворенні мікротрубочок цитоскелета. Крім участі в поділі ядра, клітинний центр відіграє важливу роль у формуванні еукаріотичних джгутиків і війок. Його центріолі формують базальне тіло, що лежить в основі джгутиків.
У організмів, позбавлених центріолей (наприклад, у сумчастих грибів, покритонасінних рослин), джгутики не розвиваються. Отже, для клітинного центра характерними є дві функції: участь у поділі клітини та організація цитоскелета.
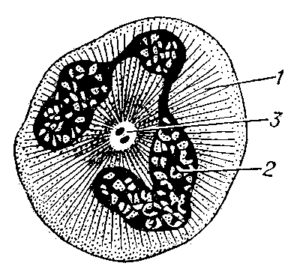 1-цитоплазма, 2-ядро, 3-клітинний центр лейкоцита
1-цитоплазма, 2-ядро, 3-клітинний центр лейкоцита





